سندرمهای بارتر و گیتلمن

یک پیشرفت عمده در مطالعه بر روی اختلالات ارثی سندرمهای دفعکنندهٔ نمک، اثبات این نکته بود که سندرمهای بارتر (Bartter ‘ssyndrome) و گیتلمن (Gitelman ‘ssyndrome)، ناشی از جهش در پروتئینهای انتقالدهندهٔ یونی خاص هستند. این پروتئینها توسط سلولهای نفرون دیستال بیان میشوند. اختلال عملکرد همانتقالدهندهٔ سدیم ـ کلر (NCCT) حساس به تیازید در سندرم گیتلمن، و اختلال در همانتقالدهندهٔ سدیم ـ پتاسیم ـ کلر (NKCC2) حساس به بومتانید در سندرم بارتر، موجب اتلاف نمک، کاهش حجم خارج سلولی، هیپرآلدسترونیسم ثانویه و هیپوکالمی میشود.
| جدول 6 ـ 28. انواع سندرم بارتر | |||||
| نوع 1 | نوع 2 | نوع 3 | نوع 4 | نوع 5 | |
| نام ژن | SLCI2AI | KCNJI | CLCNKB | BSND | CASR |
| نام پروتئین | NKCC2 | ROMK | CLCNKB | Barttin | CaR |
| علائم عمده | پلیآوری، هیپوکلسمی | پلیآوری، هیپوکالسمی | متغیر | پلیآوری، هیپوکلسمی | ـ |
| تشنج | دهیدراتاسیون | ـ | خفیف تا شدید | + کری | ـ |
| Ca2+ ادرار | ↑ | ↑ | ↑ | ↑ | ↑ |
| Mg2+ ادرار | ↓ | ↓ | ↓ | ↓ | ↓ |
| نفروکلسینوز | +++ | +++ | -+ | – | +++ |
| Ca2+ = کلسیم؛ Mg2+ = منیزیم | |||||
مشخصههای 5 سندرم مختلف بارتر و سندرمهای گیتلمن، در جداول 6 ـ 28 و 7 ـ 28 نشان داده شده است. به طور خلاصه، سندرم بارتر از نظر ژنتیکی یک بیماری ناهمگن است. بر پایهٔ مطالعات ژنتیک مولکولی، 5 زیگونهٔ مختلف از سندرم بارتر وجود دارند. نوع I یا سندرم بارتر نوزادی، ناشی از جهشهای کاهش عملکرد در همانتقالدهندهٔ سدیم ـ پتاسیم ـ کلر NKCC2 است. NKCC2 توسط ژن SCL12A1 واقع بر کروموزوم 15 رمزگذاری میشود. این همانتقالدهندهٔ سدیم ـ پتاسیم ـ کلر، بر غشای رأسی سلولهای شاخهٔ ضخیم صعودی قوس هنله بیان میشود و در حالت طبیعی، مسئول حدود 30% کل بازجذب سدیم پالایش شده توسط گلومرول است. بیماران مبتلا به این سندرم، در ابتدای زندگی دچار یک اختلال سیستمیک شدید به صورت اتلاف قابلملاحظهٔ سدیم و پتاسیم، پلی هیدرآمنیوس و هیپرکلسیوری شدید و نفروکلسینوز میشوند. ساخت و دفع پروستاگلاندین، به طور قابل ملاحظهای افزایش مییابد و ممکن است مسئول اکثر علائم سیستمیک باشد.
| جدول 7 ـ 28. پاتوفیزیولوژی شناخته شدهٔ سندرم گیتلمن |
| جهش کاهندهٔ عملکرد: NCCT اتلاف کلرید سدیم هیپرآلدوسترونیسم ثانویه: اتلاف پتاسیم هیپرپلاریزاسیون سلولی ثانویه به کاهش ورود Cl– ↑ ورود ECaC از غشای رأسی ↑ افزایش تعویض Ca/Na در غشای قاعدهای ـ جانبی اثر خالص: هیپوکلسیوری اتلاف Mg: مکانیسم نامشخص |
سندرم بارتر نوع II، ناشی از جه کاهش عملکرد در ژن KCNJ1 واقع بر کروموزوم 11 است. این ژن، کانال پتاسیم وابسته به ولتاژ یکسوکننده به داخل (inward rectifier voltage – dependent potassium channel) ROMK را رمزگذاری میکند. کانال پتاسیم ROMK، در غشای رأسی شاخهٔ ضخیم صعودی قوس هنله قرار دارد ولی در نجرای جمعکنندهٔ قشری نیز بیان میشود. در شاخهٔ ضخیم صعودی قوس هنله، جریان پتاسیم از داخل این کانال به درون لولهٔ کلیه برای فعالیت NKCC2 ضروری است که محتاج تأمین پتاسیم داخل مجرایی کافی میباشد. در مجرای جمعکنندهٔ قشری، این کانال در دفع پتاسیم موجود در رژیم غذایی نیز دخیل است. در سندرم بارتر نوع II، یک کانال ROMK غیرطبیعی، منجر به اختلال عملکرد همانتقالدهندهٔ NKCC2 و درنتیجه، اتلاف نمک، جریان بالای لولهای و اتلاف دیستال پتاسیم میشود. سندرم بارتر نوع III، توسط جهشهای کاهش عملکرد ژن CLCNKB واقع بر روی کروموزوم 1 رخ میدهد. این ژن، پروتئین کانال کلر CLC-Kb را رمزگذاری میکند. پروتئین CLC-Kb، در غشای قاعدهای ـ جانبی شاخهٔ ضخیم صعودی قوس هنله رمزگذاری میشود و مسئول بازجذب کلریدسدیم در این قسمت است.
اتلاف نمک از راه کلیه در سندرم بارتر نوع III، از انواع I و II خفیفتر است. اخیرآً، مشخص شده که سندرم بارتر نوع IV، در اثر جهش کاهندهٔ عملکرد ژن BSND (سندرم بارتر و کری حسی ـ عصبی)، واقع بر کروموزوم 1p31 ایجاد میشود. ژن BSND، بارتین (barttin) را رمزگذاری میکند. این پروتئین در غشای قاعدهای ـ جانبی شاخهٔ ضخیم صعودی قوس هنله بیان میشود. بارتین، زیرواحد بتای کانال کلر C1C-Kb است و برای رسیدن C1CKb بع غشای پلاسمایی، لازم میباشد. این پروتئین در حلزون گوش (cochlea) همراه ابا کانالهای کلر C1CKa و C1CKb قرار دارد. این بیماران، همچنین دچار اتلاف سدیم و پتاسیم و اختلال عملکرد حلزون و کری میشوند. اخیراً آلکالوز هیپوکالمیک ناشی از سندرم بارتر نوع V، در بیماران مبتلا به هیپوکلسمی اتوزوم غالب یافت شده است. در این بیماری، هیپوکلسمی به جهشهای افزایندهٔ عملکرد در گیرندهٔ حسکنندهٔ کلسیم (CaSR) مربوط میشود. CaSR، در غشای قاعدهای جانبی شاخهٔ ضخیم صعودی قوس هنله به میزان زیادی بیان میشود و تصور میشود که نقش مهاری مهمی در تنظیم انتقال خلال سلولی (transcellular) سدیم، کلر و کلسیم دارد. فعال شدن CaSR قاعدهای جانبی در شاخهٔ ضخیم صعودی قوس هنله، فعالیت کامال پتاسیم رأسی را کاهش میدهد که باعث ایجاد سندرمی مشابه بارتر میگردد. همچنین انتظار میرود فعال شدن ژنتیکی CaSR توسط این جهشها، از طریق مهار تولید اختلاف پتانسیل مثبت مجرایی که منجر به انتقال جنب سلولی کلسیم در شاخهٔ ضخیم صعودی قوس هنله میشود، دفع ادراری کلسیم را افزایش دهد.
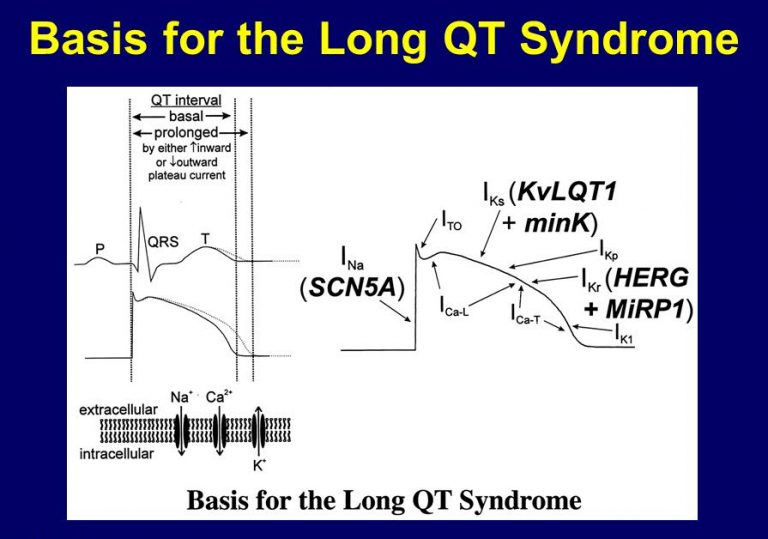
تا امروز، به نظر میرسد سندرم گیتلمن از نظر مولکولی همگن باشد. اگرچه جهش کاهندهٔ عملکرد در ژن SLC12A3 واقع بر کروموزوم 16q13 به عنوان یکی از علل شناخته میشود، بررسیهای ژنتیک اخیر بر روی سندرم گوردون (Gordon ‘ssyndrome) که مشخصات بالینی آن برعکس سندرم گیتلمن است، احتمال اینکه جهشهای آینهای مشابه (و احتمالاً بیش از یک جهش) مسئول سندرم گیتلمن باشند را مطرح میسازد. ژن SLC12A3، همانتقالدهندهٔ کلیوی سدیم ـ کلر حساس به تیازید NCCT را رمزگذاری میکند. NCCT، مسئول بازجذب سدیم در لولهٔ دور است و حدود 7% کل سدیم پالایش شده را بازجذب میکند. اگرچه سندرم گیتلمن از سندرم بارتر خفیفتر است، بیماران دچار مشکلات قابل توجه ناشی از علائم عضلانی، ضعف، و افزایش خطر آریتمیهای قلبی (در موارد طولانی شدن فاصلهٔ QT) میشوند. علیرغم افزایش فعالیت رنین پلاسما، دفع کلیوی پروستاگلاندین بالا نمیرود و این خصوصیت، یکی دیگر از تفاوتهای سندرم گیتلمن نسبت به سندرم بارتر است.
یک تفاوت فنوتیپی عمده بینن سندرم بارتر و سندرم گیتلمن، دفع کلسیم از راه ادرار است. عقیده بر این است که هیپرکلسیوری مربوط به سندرم بارتر، عمدتاً ناشی از اختلال عملکرد سلولهای شاخهٔ ضخیم صعودی لولهٔ هنله است. جذب کلسیم که فرآیندی غیرفعال و جنب سلولی است، در اثر ولتاژ مثبت خلال اپی تلیومی مجرا انجام میگیرد. این ولتاژ، توسط همانتقالدهندهٔ NKCC2 و بازیافت مجرای K+ تولید میشود. در صورت کاهش یا مهار همانتقالی NKCC2 توسط داروهای مدر مؤثر بر قوس هنله یا ناهنجاریهای ژنتیک، از مثبت بودن مجرا کاسته میشود و درنتیجه بازجذب کلسیم، کاهش مییابد. چیزی که به طور کامل مشخص نیست، این است که چرا کاهش مثبت بودن مجرا منجر به افزایش دفع منیزیم نمیشود. علاوه بر این مکانیسم، افزایش رسیدن NaCl به لولهٔ دیستال ناشی از اختلال عملکرد شاخهٔ ضخیم صعودی، غلظت داخل سلولی کلر را بالا میبرد که به نوبهٔ خود، کانال کلسیم رأسی سلولهای لولهٔ پیچیدهٔ دور را مهار میکند و درنتیجه، منجر به احتباس بیتر کلسیم و سنگ کلیه میود. برعکس بیماران مبتلا به سندرم بارتر، بیماران مبتلا به سندرم گیتلمن بلااستثنا دچار هیپوکلسیوری هستند.
مکانیسم هیپوکلسیوری سندرم گیتلمن، مشابه اثرات بالینی مفید داروهای مدر مؤثر بر لولهٔ پیچیدهٔ دور (تیازیدها و داروهای دیگر) در کاهش دفع ادراری کلسیم است. مکانیسم هیپوکاسیوری در سندرم گیتلمن به خوبی شناخته شده است. ابتدا کاهش خفیف در حجم ECF منجر به افزایش بازجذب کلسیم از لولهٔ نزدیک میشود و سپس، کاهش ورود NaCl به سلولهای لولهٔ پیچیده دور، انتقال خلال اپیتلیومی کلسیم را تحریک میکند. وقتی وارد شدن Na و Cl از غشای رأسی سلولهای لولهٔ پیچیدهٔ دور مهار میشود (چه به دلیل استفاده از داروهای مدر و چه ناشی از بیماری ژنتیک)، غلظت دخل سلولی کلر کاهش مییابد. فعالیت کمتر Cl داخل سلولی، منجر به هیپرپلاریزه شدن سلول میشود و ورود کلسیم به داخل سلولها از طریق کانالهای خاص ورود کلسیم ECaC و CaT2 رأسی که بر روی سلولهای لولهٔ پیچیده دور بیان میشوند را فعال میکند. از آ«جا که حرکت کلسیم از مجرا به سلول باید به نحوی جبران شود، افزایش ورود کلسیم به سلولهای لولهٔ پیچیدهٔ دور، موجب تحریک خارج شدن سدیم از طریق معاوضهکنندهٔ Ca+/Na+ و Ca-ATPase واقع در غشای قاعدهای ـ جانبی میشود. بنابراین، حاصل امر، به وجود آمدن هیپوکلسیوری است.
گرچه پاتوژنز اختلالات کلسیم در سندرمهای بارتر و گیتلمن نسبتاً واضح است اما فقط اخیراً پاتوژنز اختلالات منیزیم همراه با این سندرمها روشن شده است. سندرم گیتلمن یا هیپومنیزیمی شدید همراه است درحالی که سندرم بارتر اینگونه نیست. مشاهدات اخیر از اختلالات ژنتیکی هیپومنیزیمی و نیز مشاهدات بالینی در بیمارانی که توسط آنتیبادیهای ضدعامل رشد اپیدرمی تحت شیمیدرمانی قرار میگرفتند به روشن ساختن مکانیسمهای مولکولی دخیل در نقل و انتقال منیزیم کمک کرده است. منیزیم در سرتاسر طول نفرون جذب میشود اما جایگاه اصلی بازجذب آن در طول لولهٔ دور میباشد. منیزیم به همراه کلسیم از طریق یک مسیر جنب سلولی با خاصیت گزینشی نسبت به بار الکتریکی در شاخهٔ ضخیم صعودی قوس هنلهٔ مدولاری و قشری بازجذب میشود.
جهشهایی در کلائودین (claudin) 16 (که پاراسلین نیز نامیده میشود) و کلائودوین 19 باعث هیپومنیزیمی شدید و نفروکلسینوز میشود. در لولهٔ پیچیدهٔ دور یک کانال ویژه به نام گیرندهٔ گذرای پتانسیل ملاستاتین (transient receptor potential melastatin) زیرخانوادهٔ 6 (TRMP6) واسطهٔ بازجذب منیزیم است. جهش در ژن عامل رشد اپیدرمی که در لولهٔ دور بیان میشود باعث بروز هیپومنیزیمی میگردد؛ آنتیبادیهای ضدعامل رشد اپیدرمی نیز هیپومنیزیمی ایجاد میکنند. مشاهدهٔ اخیر مطرحکننده یک نقش تنظیمی برای عامل رشد اپیدرمی در بازجذب منیزیم است. غیرفعال کردن همانتقالدهندهٔ سدیم ـ پتاسیم ـ کلر حساس به تیازید و مهار این ناقل با تیازیدها موجب هیپومنیزیمی و کاهش بیان TRPM6 میشود. درنتیجه کاهش بیان کانال TRPM6 محتملترین توجیه هیپومنیزیمی مشاهده شده در سندرم گیتلمن است.
مطالب مرتبط